Г-Я-взаимодействие.
Физиология человека и животных

Хелперы также делятся на две группы: 1-го типа (7х-1) и 2-го типа (Тх-2). Именно Тх-2 взаимодействуют с 5-клетками, запуская развитие новых клопов и выброс в кровь дополнительных антител. Тх-1 усиливают реакцию иммунной системы иным способом: после встречи со «своим» антигеном они начинают выделение в межклеточную среду цитокина интерферона-у (рис. 5.9). Это соединение вызывает ярко выраженную… Читать ещё >
Г-Я-взаимодействие. Физиология человека и животных (реферат, курсовая, диплом, контрольная)
й-л имфоциты также относятся к группе АПК, и на их поверхности можно обнаружить связанные с белками ГКГС антигены. При этом первичный захват антигена происходит в результате связывания с расположенным на наружной мембране й-лимфоцита антителом. После связывания антитело вместе с антигеном погружается в цитоплазму, где происходит сначала расщепление чужеродной молекулы, а затем размещение ее фрагментов на белках ГКГС (рис. 5.8).
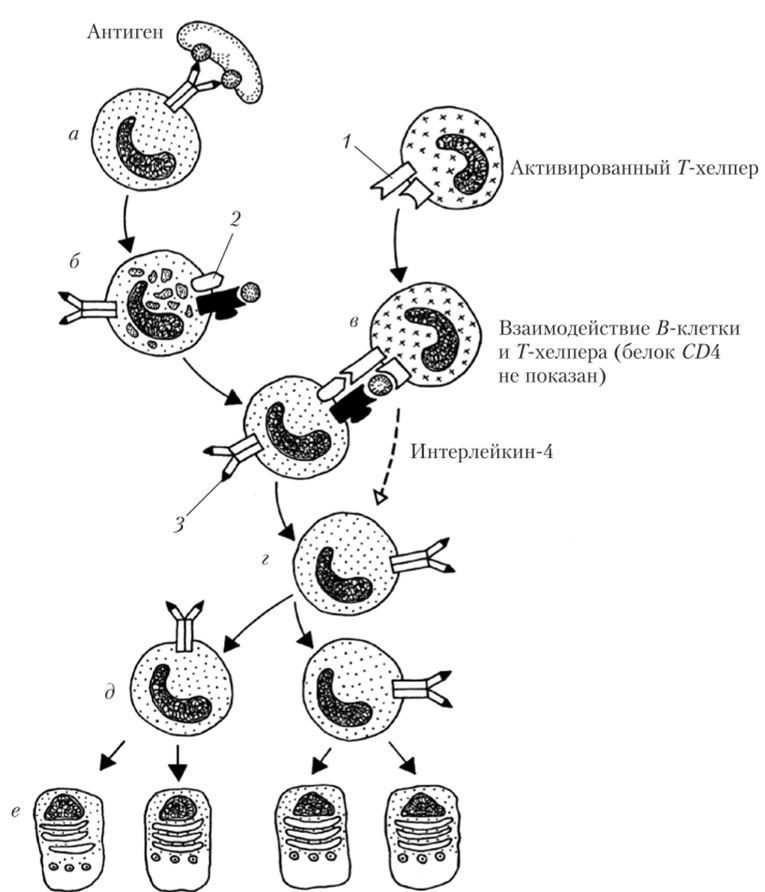
Рис. 5.8. После присоединения антигена в антителу (а) 5-лимфоцит презентирует его эпитоп на своей поверхности (б). При встрече с активированным Г-хелпером, рецептор которого настроен на тот же эпитоп (в), происходит активация 5-клетки с помощью интерлейкина-4 (г) и размножение соответствующего клона (д) с образованием плазматических клеток (е):
1 — белок CDA0L; 2 — белок CD АО; 3 — антитело А5-клетки; остальные обозначения — как на предыдущем рисунке
Таким образом, эпитоп, на который специфически настроен определенный клон 5-лимфоцитов, при появлении в малой концентрации не вызывает прямой активации S-клеток. Однако он оказывается презентирован на поверхности 5-лимфоцитов и способен к взаимодействию с Г-рецепторами.
Активированные Г-хелперы проявляют стремление к контакту с 5-клетками. Их рецепторы «ищут» на мембране 5-лимфоцитов соответствующий эпитоп и, если находят, то присоединяются к нему, формируя так называемые иммунологические синапсы (это чаще всего происходит внутри лимфатического узла).
В иммунологическом синапсе рецепторы и CZM-белки Г-клетки контактируют с эпитопами антигена и белками ГКГС 5-лимфоцита. Кроме того, CDA0L взаимодействует с белком CDA0 (см. рис. 5.8). В результате 5-лимфоцит получает разрешение делиться и формировать клон с последующим развитием плазматических клеток. Кроме того, Г-хелпер, который встретил «свою» 5-клетку, начинает выделять в окружающую среду интерлейкин-4 — цитокин, значительно ускоряющий деление 5-лимфоцитов.
Подчеркнем еще раз, что описанный путь передачи антигенного сигнала от АПК к Г-хелперам и далее к 5-лимфоцитам обладает свойствами мощного усилительного механизма, позволяющего запустить продукцию антител при низкой концентрации антигена в организме.
Приведем несколько дополнительных замечаний. Белки класса ГКГС делятся на две группы: тип I и тин II (ГКГС-I и ГКГС-П). Выше речь шла только о ГКГС-П, которые свойственны АПК и презентируют захваченные из межклеточной среды антигены. В противоположность этому ГКГС-I присущи всем клеткам и служат для презентации фрагментов собственных белков организма (см. рис. 5.5, слева). Контакт с молекулами ГКГС-I и связанными с ними эпитопами наших клеток осуществляют Т-киллеры, которые не должны встретить на «тарелочках» чужеродные молекулы. Если такая встреча все же произошла, это означает, что клетка, «отчитавшаяся» с помощью ГКГС-I перед иммунной системой, заражена вирусом либо мутировала. В обоих случаях запускается механизм се уничтожения (см. ниже). Отметим также, что белки ГКГС характеризуются гигантским разнообразием аллелей. В связи с этим полученный от родителей набор молекул ГКГС уникален для каждого человека. Именно эта неповторимость является основной причиной отторжения пересаженных органов, поскольку иммунная система реагирует па фрагменты белков ГКГС донора как на чужеродные антигены.
7-хелперы также делятся на две группы: 1-го типа (7х-1) и 2-го типа (Тх-2). Именно Тх-2 взаимодействуют с 5-клетками, запуская развитие новых клопов и выброс в кровь дополнительных антител. Тх-1 усиливают реакцию иммунной системы иным способом: после встречи со «своим» антигеном они начинают выделение в межклеточную среду цитокина интерферона-у (рис. 5.9). Это соединение вызывает ярко выраженную активацию макрофагов и фагоцитов крови. В результате инфекция может быть устранена средствами врожденного клеточного иммунитета (без включения «обучения»).
В момент встречи с АПК и «своим» антигеном путь, по которому пойдет развитие Г-хелпера, еще не определен. Выбор, стать Тх- или Тх-2, производится каждой клеткой клона отдельно — в зависимости от так называемого цитокинового окружения (соотношения концентрации разных цитокинов в конкретной части организма).
С учетом всего сказанного можно выделить три «линии обороны», характерные для иммунной системы. Они последовательно срабатывают, если инфекционный процесс нарастает, причем вторая из них конкурирует с третьей (аналогичным образом, например, при реализации оборонительного поведения конкурируют реакции страха и ярости — «беги либо дерись»).
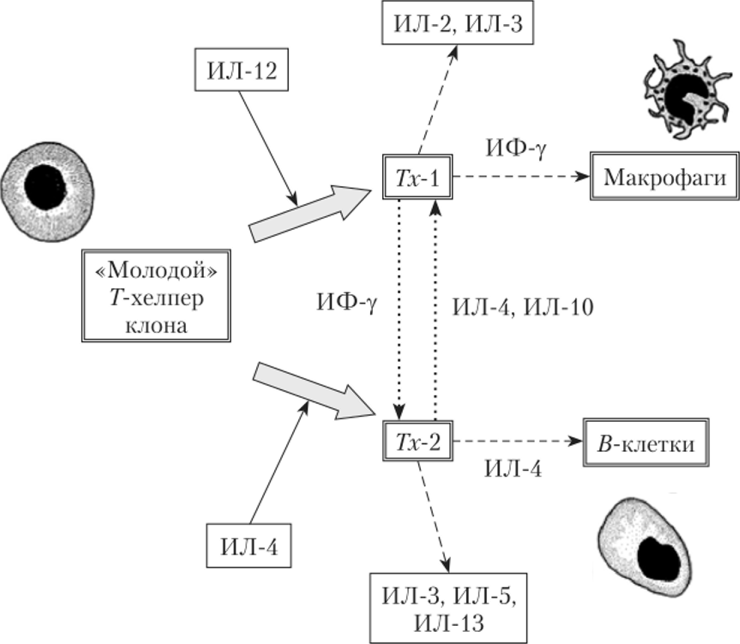
Рис. 5.9. Выбор пути развития Г-хелпером:
превращение «молодого» Г-хелпера размножающегося клона в Тх- либо Тх-2
происходит иод действием ряда интерлейкинов (сплошные стрелки). Далее Тх- и Тх-2 выделяют цитокины, активирующие другие клетки иммунной системы (прерывистые стрелки), а также цитокины, с помощью которых Тх- и Тх-2 конкурируют друг с другом (пунктирные стрелки).
- 1. На антигены, по той или иной причине «знакомые» иммунной системе, реагируют имеющиеся в организме й-клетки памяти и соответствующие антитела, а также элементы врожденного иммунитета (комплемент, макрофаги). Активность составляющих врожденного иммунитета значительно усиливается при предварительной онсонизации антигенов и (или) их источников антителами.
- 2. Если инфекционный агент не устранен и в организме начинает нарастать концентрация новых для него антигенов, активируются дополнительные клоны Г-хелперов, причем основная часть клеток становится Тх-. Это происходит под влиянием интерлейкина-12 — фактора, выделяемого макрофагами (рис. 5.9, вверху). Тх- начинают выброс интерферона-у, который дополнительно «подхлестывает» макрофаги и фагоциты крови, повышает их чувствительность к чужеродным молекулам. Возможно, этого окажется достаточно для устранения инфекции.
- 3. Если содержание новых антигенов в организме продолжает увеличиваться, то становится более значимым влияние на Г-лимфоциты интерлейкина-4 (рис. 5.9, внизу). Он, с одной стороны, тормозит продукцию интерферона-у, с другой — способствует выбору пути Тх-2. Источником интерлейкина-4 служат уже активированные Тх-2, а также киллерные и тучные клетки. Развитие новых клонов Тх-2 приводит к включению дополнительных клонов й-лимфоцитов, обогащению набора присутствующих в крови антител, повышению специфичности и интенсивности реагирования иммунной системы па источник антигенов. Тх-2, кроме интерлейкина-4, продуцируют ряд других цитокинов. В их числе — интерлейкин-10, тормозящий активность Тх-1. В свою очередь, Тх-l ослабляют деятельность Тх-2 с помощью интерферона-у.
Последовательность включения перечисленных механизмов показывает, что иммунная система стремится (при возникновении инфекции) ограничиться теми средствами, которые уже имеются, и активация новых клонов происходит лишь «в крайнем случае». Здесь видна еще одна аналогия с работой мозга: НС, особенно взрослая и адаптировавшаяся к окружающей среде, также зачастую неохотно учится, пытаясь обойтись теми навыками и стереотипами, которые сформированы ранее.