Общая характеристика нервной системы позвоночных

Современные позвоночные представляют собой конечные этапы широкой и многоступенчатой адаптивной радиации. Они довольно рано начали утверждаться на высших ступенях трофических пирамид, что давало каждой отдельной группе определенные преимущества, но при этом обусловливало и ряд сложностей, например в добывании пищи. Экологическая дифференциация позвоночных, сопровождающаяся у разных форм… Читать ещё >
Общая характеристика нервной системы позвоночных (реферат, курсовая, диплом, контрольная)
Современные позвоночные представляют собой конечные этапы широкой и многоступенчатой адаптивной радиации. Они довольно рано начали утверждаться на высших ступенях трофических пирамид, что давало каждой отдельной группе определенные преимущества, но при этом обусловливало и ряд сложностей, например в добывании пищи. Экологическая дифференциация позвоночных, сопровождающаяся у разных форм интенсификацией функций органов, определялась прогрессивным усложнением среды их обитания. Таким образом усилился отбор по способности адекватно отвечать на весьма разнообразные изменения среды, что обеспечивалось прежде всего развитием нервной системы, поскольку частные приспособления в отличие от преобразований ЦНС не имеют столь универсального значения в изменчивой и сложной обстановке. При этом процессы усложнения и дифференциации нервной системы и ее структурных признаков, протекая в сходных условиях, в филогенетически далеких ветвях позвоночных могли приводить к формированию в ЦНС функционально сходных, но отличающихся по своему происхождению структур (конвергенция). Процессы же усложнения среды обитания при наличии общей наследственной основы у близкородственных форм могли в еще большей степени определить однонаправленные преобразования в ЦНС. Параллельные преобразования в нервной системе распространены намного шире, хотя в близкородственных группах трудно выявить результаты таких параллельных, независимых изменений.
До недавнего времени характеристику отдельных групп позвоночных проводили на примере небольшого числа их типичных представителей — «модельных» форм, которые не всегда отражают многообразие признаков конкретного уровня организации. При обилии случаев независимого усложнения ЦНС в разных ветвях позвоночных наиболее подходящими объектами для выявления былых эволюционных стадий среди современных форм являются экологически консервативные реликты, т. е. формы, сохраняющие древнюю организацию и обитающие в среде, по абиотическим и, особенно, биотическим обстоятельствам мало отличающейся от среды обитания их древних предков. При этом для нахождения различий в функциональных генотипических свойствах, связанных с особенностями условий среды, следует сравнивать систематически близкие виды животных. Для нахождения же различий, связанных с происхождением, необходимо сравнивать животных, обитающих в одинаковых условиях. Однако при анализе получаемых данных должна учитываться возможность весьма значительной неравномерности хода эволюции, а также определенная «мозаичность» преобразования разных признаков структур, в том числе ЦНС, функций в линии предок-потомок. Формирование признаков проходило неравномерно и нередко параллельно, независимо даже в близкородственных группах. При этом «принцип мозаичности» может проявляться как в сохранении у одних представителей группы тех или иных черт древнего, «анцестрального», состояния, так и в возникновении у других широкого спектра признаков иных уровней организации (часто более высоких, чем у типичных «модельных» форм), нередко как бы преадаптирующих свойства вышестоящих групп и типов[1].
В связи с этим, прежде чем переходить непосредственно к рассмотрению вопросов эволюции нервной системы позвоночных, необходимо краткое изложение современных представлений о филогенетическом развитии и систематике позвоночных.
Становление типа хордовых, Chordata, шло в докембрии, повидимому, в период, когда примитивные оболочники (подтип Tunicata), обладающие личинкой, переходили к неотении и потере прикрепленной стадии. Бесчерепные (подтип Acrania), произошедшие от таких форм, в ходе морфофизиологического развития дали начало позвоночным (подтип Vertebrata) (рис. 1).
Позвоночные животные разделяются на две крупные ветви: бесчелюстные (Agnatha), которые представлены сейчас одним классом Круглоротые (Cyclostomata) и челюстноротыми (Gnathostomata). Исходной группой для обеих ветвей были телодонты (подкласс Thelodonti класса Pteraspidomorphi) — некрупные беспанцирные формы, обитавшие в прибрежных участках морей и в лиманах. Появление у бесчелюстных панциря могло сопровождаться некоторым упрощением их связей со средой. Примитивные костнощитковые бесчелюстные (в основном относящиеся к классу Cephaloaspidomorphi) были придонными, проходными обитателями дельтовых водостоков и лиманов. Они, видимо, обладали развитыми системами как электропродукции, так и электрорецепции.
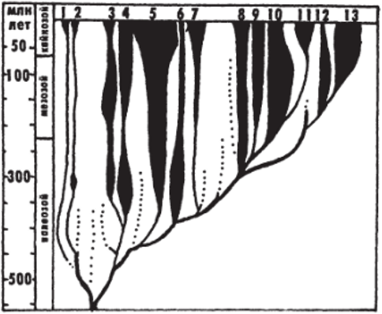
Рис. I
Схема эволюции основных классов позвоночных (Fite, 1985).
1 — бесчерепные; 2 — круглоротыс; 3—7 — рыбы: цельноголовые (.?), пластиножаберные (4), лучеперые и многоперые (5), сарколторигин (в), двоякодышащие (7); 8 — амфибии; 0−11 — рептилии: черепахи (О), чешуйчатые (10), архозавры (//); 12. 13 — млекопитающие: первоявери (однопроходные) (12), звери (сумчатые и плацентарные) (13)
и лишь после достижения максимальных размеров тела приобретали панцирную защиту. Головной мозг первичных бесчелюстных был вытянут в одной плоскости, обонятельные лопасти не обособлены от переднего мозга, хабенулы промежуточного мозга асимметричны, мозжечок уже достаточно хорошо развит (рис. 2, а). Задний мозг был связан с октаво-латеральной системой, средний — со зрительной, передний — с обонятельной. Таким образом, уже на этой стадии эволюции головной мозг дифференцировался на ряд отделов, возникновение и развитие которых, как полагают, теснейшим образом связано с развитием основных систем дистантных анализаторов.
Круглоротые (Cyclostomata) произошли от нектонных бесщитковых бесчелюстных подкласса Anaspida, некоторые формы которых, имевшие лишь кератиновые чешуи, приспосабливались к использованию тел погибших панцирных бесчелюстных как временных убежищ и источников пищи. Сформировавшиеся таким образом миноги (подкласс Petromyzones) переходили к эктопаразитизму и полухищничеству в пресных и солоноватых водоемах, а миксины (подкласс Myxini) — к особой форме хищничества в море. Разделение двух ветвей круглоротых произошло еще до начала карбона. На этом длительном пути эволюции от Anapsida к типичным миногам и, особенно, к миксинам могло произойти некоторое вторичное упрощение в строении и функционировании их ЦНС. Тем не менее круглоротые являются первым классом позвоночных, у которых сформированы пять основных отделов головного мозга (продолговатый, задний, средний, промежуточный и конечный), характерные и для всех остальных позвоночных, включая человека.
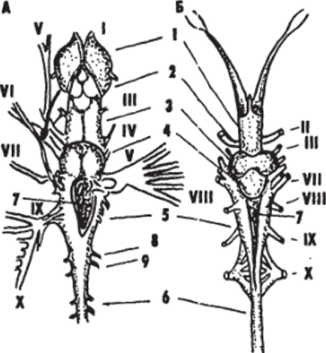
Рис. 2.
Головкой мозг древней бесчелюстной рыбы Cephalaspis (Л) и древней хрящевой акулоподобной рыбы Arthrodira (Б) (Стеншио, 1925 — 1927).
I — 6 — отделы головного мозга: передний (1), промежуточный (2), средний (Я), мозжечок (4), продолговатый (5), спинной (0); 7 IV мозговой желудочек; 8, 9 — брюшные (8) и спинные (9) спинномозговые корешки; I — X — черепно-мозговые нервы Преобразование первых жаберных дуг в челюстной аппарат, служащий для активного схватывания пищи, и развитие парных плавников дали первым челюстноротым (Protognathostomata) значительные эволюционные преимущества перед бесчелюстными. Способность к активному питанию резко повысила их жизнедеятельность и позволила завоевать водную среду, развившись в самый многочисленный и разнообразный по жизненным формам надкласс Рыбы (Pisces).
Одно из крупных подразделений рыб включает классы Панцирные рыбы (Placodermi — ныне вымершие), и класс Хрящевые рыбы (Chondrichthyes). Появившись в начале девонского периода, хрящевые рыбы в большей степени, чем панцирные, сохранили облик и основные особенности организации предковых форм. Пластиножаберные (подкласс Elasmobranchii), представляющие собой крупнейшую группу хрящевых рыб, в палеозое были весьма многочисленны в солоноватых водах морей и лиманов, в кайнозое придерживались, в основном, морей. Они разделяются на акул и скатов, которые представляют собой не ветви, а ступени эволюции. Эласмобранхи со скатообразным телом не раз возникали от акулоподобных предков. В настоящее время консервативными реликтами палеозоя являются плащеносные акулы, с юры известны морские ангелы и гребнезубые акулы, с мела — пилоносы, пилы-рыбы, разнозубые акулы. Особую, в целом прогрессивную, группу пластиножаберных рыб составляют ламноидные акулы, кархарины и некоторые ветви скатов. Они играют значительную роль в экосистемах морей, занимая вершины трофических пирамид. Современные Хрящевые рыбы — процветающая и ныне группа рыб, заселившая обширные районы Мирового океана и некоторые пресноводные водоемы и насчитывающая в своем составе около 50 семейств и несколько сот видов акул, скатов и химер.
Как показала реконструкция (по эндокранам) головного мозга древних девонских акулоподобных Arthrodira из группы Macropetalich;
thys, они обладали сложным мозгом с типичным разделением на пять отделов (рис. 2. б). Крупные обонятельные лопасти мощными обонятельными нервами соединялись с обонятельными луковицами, расположенными на вентральной стороне конечного мозга. Средний мозг почти в два раза превышал размеры переднего. Крупный мозжечок почти прикрывал крышу среднего мозга и часть ромбовидной ямки продолговатого мозга. Хорошо развитыми являлись черепно-мозговые нервы V — X пар с наиболее мощным блуждающим нервом.
Цельноголовые (подкласс Holocephali) по ряду черт организации занимают промежуточное положение между пластиножаберными и панцирными рыбами. В мелу они обитали в прибрежных водах морей, однако в дальнейшем, вследствие развития костистых рыб, оказались оттесненными в более простую среду обитания — на средние и большие глубины.
Второе крупное подразделение рыб включает преимущественно солоноватоводных и пресноводных челюстножаберных рыб (класс Acanthodii — ныне вымершие) и возникших от них в девоне костных рыб[2]. Предки костных рыб очень рано разделились на подклассы лучеперых рыб (Actinopterygii), многоперов (Polypterigii) и мясистолопастных (Sarcopterygii). Древние лучеперые рыбы из надотряда палеонисков (Palaeonisci) уже в палеозое очень широко дивергировали. Так, от одного из стволов палеонисков серией ветвей возникли цельнокостные рыбы (надотряд Holostei), обитавшие в солоноватых и пресных водах мезозоя. Ныне эта группа представлена лишь двумя семействами консервативных реликтов мелового периода — панцирными щуками и ильными рыбами. За счет параллельных преобразований не менее чем трех ветвей древних голостей постепенно сформировалась группа костистых рыб (надотряд Теleoste;). Появившись в мелких лиманных водах палеозоя, они постепенно замещали здесь цельнокостных, а к концу раннего мела распространились в морях и пресных водах. По-видимому, от особого ствола палеонисков, развиваясь по пути фетализации (задержки признаков ранних стадий онтогенеза до более поздних стадий индивидуального развития), возникли хрящевые ганоиды (надотряд Chondrostei), которые уже в середине мела были разделены на ветви осетровых рыб и веслоносов. Многоперы, видимо, существуют в малоизменившейся среде с палеозоя. Среди мясистолопастных рыб из надотряда кистеперых (Crossopterygii) было несколько стволов. Хищники-засадчики солоноватых и пресных вод палеозоя, объединяемые под названием Rhipidistia, дали начало наземным позвоночным или четвероногим (надкласс Tetrapoda). Группу Rhipidistia нередко противопоставляют целакантам (отряд Coelacanthiformes), которые еще в девоне, не успев даже приобрести некоторые черты организации, характерные для Rhipidistia, ушли в глубокие морские воды, в которых они, очень медленно изменяясь, дожили до современности в качестве «живого реликта» — латимерии.
Современные пресноводные двоякодышащие рыбы (надотряд Dipnoi) обитают, в основном, в среде той же сложности, что и их древние предки, отделившиеся от форм, общих с кистеперыми. Две современные ветви — австралийская и африкано-южноамериканская развиваются самостоятельно по меньшей мере с конца палеозоя. Такие консервативные поведенческие адаптации, как образование «кокона» при высыхании водоемов, впадение в спячку, зарывание в грунт также, видимо, сформировались еще в конце палеозоя в среднем и позднем девоне. Отметим, что по современным представлениям двоякодышащие рыбы не имеют отношения к происхождению наземных позвоночных.
Исходной группой костных рыб, от которых произошли первые наземные позвоночные (Tetrapoda), являются древние кистеперые рыбы из группы рипидистий (Rhipidistia). Как сейчас признается палеонтологами, климатические условия в девонском периоде (времени выхода позвоночных на сушу) были очень изменчивы. Это способствовало значительной адаптивной радиации саркоптеригий. Одни рипидистии активно осваивали пресноводные водоемы, другие переходили к жизни в морских лагунах, где, подвергаясь периодическим опреснениям, по-видимому, смогли приспособиться к полуводному образу жизни, приобретя, таким образом, свойства, послужившие в дальнейшем предпосылками выхода на сушу.
Необходимо подчеркнуть, что формирование многих признаков, характерных для амфибий, происходило у их предков еще в водной или полуводной среде. Они оказались способными относительно быстро занимать случайно освобождающиеся места в экосистемах водоемов, замещая кистеперых в их исходной водной и, отчасти, полуводной среде. Таким образом, приспособление к крайне необычным для рыб условиям жизни привело в эволюции позвоночных к выработке адаптаций, обеспечивших освоение новой для них наземной среды. Приобретение этих адаптаций происходило среди филетически различных групп рипидистий, и ряд «полуамфибий» так и остались слепыми ветвями эволюции. Строгой и единственной линии, ведущей от рыб к амфибиям, не существовало. Современные амфибии, составляющие надотряд Lissamphibia, представлены тремя отрядами: хвостатыми (Urodela), бесхвостыми (Anura) и безногими (Apoda). За этими тремя группами признается монофилетическое происхождение, причем бесхвостые амфибии являются потомками ствола, более 350 млн. лет развивающегося обособленно от хвостатых и безногих. Их формирование происходило в палеозое от достаточно сильно различающихся предковых групп, обитавших на разных массивах суши, относящихся в настоящее время, соответственно, к Северному и Южному полушарию. Переход к наземному образу жизни способствовал значительной дифференцировке и усложнению как органов чувств, так и связанных с ними отделов ЦНС.
Эволюционная ступень амниот (Amniota) включает две ветви, одна из которых, зауропсидная, содержит классы Рептилии (Reptilia) и Птицы (Aves), а другая, терапсидная, включает классы Зверообразные (Theromorpha) и Млекопитающие (Mammalia). Данные палеонтологии, сравнительной морфологии и физиологии показывают, что обе эти ветви могли достичь уровня амниот вполне самостоятельно, при этом ряд важных признаков организации сформировался еще на «амфибийной* стадии развития. Усложнение и интенсификация функций органов чувств и органов движения у амниот, усложнение их поведения в новой, более разнообразной среде обитания (амниоты полностью освоили наземную, водную и воздушную среду), без сомнения связаны с усложнением ЦНС.
Среди современных рептилий относительно прямыми потомками первых рептилий котилозавров (подкласс Cotylosauria) могут рассматриваться черепахи (подкласс Testudines), стоящие во многом особняком от остальных рептилий[3]. Из современных только пре: сноводные черепахи (но не плавающие, а ходящие по дну мелких водоемов) смогли в наибольшей степени сохранить образ жизни предковых форм и уровень их морфо-функциональной организации. Двукоготные и мягкокожие черепахи также мало изменились за последние 100 млн. лет, сохранив многие из тех связей со средой, которые установились у рубежа раннего мела. Предки морских черепах отделились от общей линии почти 120 млн. лет назад, бокошейные — почти с триаса.
От мелких, подвижных насекомоядных котилозавров (хищных по способу питания) ведет свое начало другая группа рептилий — чешуйчатые (подкласс Lepidosauria). Сохранившаяся с начала мезозоя и до наших времен реликтовая форма из отряда клювоголовых (отряд Rhynchocephalia) — гаттерия обитает в довольно простой биотической среде. Широкая адаптивная радиация ящериц (отряд Lacertilia) современной фауны происходила в основном с середины мезозоя. Нынешние группы гекконовых, агамовых, сцинковых, настоящих ящериц, Барановых и других характеризуют разные и, подчас, параллельные ветви развития рептилий. Змеи (отряд Serpentes), являющиеся филогенетически самым молодым отрядом рептилий, прошли в меловом периоде стадию роющих предков, имевших конечности. На вторичность процесса утраты конечностей у змей указывает, в частности, тот факт, что у них сохраняются в рудиментарном виде вся мышечная система конечностей и соответствующие нейронные объединения спинного мозга. В группе безногих ящериц (отряд Amphisbaenia) утрата конечностей и переход к роющему образу жизни в более простой среде обитания также вторичен по отношению к их активным и подвижным ящеричным предкам.
Архозавры (подкласс Archosauria), или высшие рептилии, также возникли непосредственно от котилозавровых предков еще на «прорептилийной» стадии развития. Уже в мезозое члены этой крупной группы рептилий (типичными представителями которой были динозавры) обладали сложным поведением, некоторые из них приобрели определенный уровень теплокровности (отряд летающих ящеров Pterosauria). Единственные из оставшихся в настоящее время архозавров — крокодилы (отряд Crocodilia) являются лишь умеренно высокими по уровню организации в группе архозавров. Наибольшим числом примитивных черт организации среди современных крокодилов обладают аллигаторы и кайманы.
С позиций эволюционной нейроморфологии рептилии представляют исключительный интерес, поскольку именно в этом классе позвоночных закладываются те признаки организации ЦНС, которые дали начало развитию высших центров головного мозга птиц и млекопитающих.
В настоящее время на смену гипотезе происхождения птиц (класс Aves) от ранних архозавров (отряд Thecodontia) пришла гипотеза их возникновения от группы мелких хищных динозавров из отряда Saurischia. Последние обладали бипедией, имели относительно большой объем головного мозга и ряд других признаков сходства с птицами. Полагают, что в эволюции позвоночных было несколько попыток выйти в воздушную среду, некоторые из которых оказались тупиковыми (например Archeopteryx). Более того, одна из переходных от рептилий к птицам форм, названная Protoavis, имея возраст, более чем на 75 млн. лет превышающий возраст археоптерикса, обладает большим набором «птичьих» признаков. Расхождение основных стволов птиц на килевые и бескилевые произошло очень рано — в мелу, тогда же началось интенсивное формирование стволов надотрядного и отрядного рангов. Полагают, что современные птицы из отрядов казуаров, тинаму и сорных кур живут в среде, близкой по уровню сложности к той, в которой жили их предки в меловом периоде мезозоя.
Два главных ствола класса Млекопитающие (Mammalia) сформировались еще на «рептилийной» стадии и ныне представлены первозверями (подкласс Prototheria) и зверями (подкласс Theria). Последние в наше время включают сумчатых (инфракласс Metatheria) и плацентарных (инфракласс Eutheria) млекопитающих. Современные первозвери — утконосые и ехидновые — разошлись в своем эволюционном развитии очень рано — еще в мезозое. Они крайне уклонились по образу жизни от своих предков и наряду с сохранением древних примитивных признаков приобрели множество специфических адаптаций.
Сумчатые и плацентарные представляют собой две ветви млекопитающих, отошедшие от общей предковой группы триботерий (Tribotheria) на рубеже раннего и позднего мела. Консервативными реликтами того времени среди сумчатых являются опоссумовые. Наиболее архаичные плацентарные относились к отряду ныне вымерших млекопитающих из отряда Protheutheria, представители которого жили в конце раннего мела — палеогене. От самых ранних форм протеутерий отделились будущие неполнозубые млекопитающие и (не менее чем тремя параллельными ветвями) формы тенрекообразных, ежеобразных и землеройкообразных, объединяемые в отряд насекомоядных (Insectivora)[4]. В конце мела именно от протеутерий отделились ветви, давшие первых приматов, рукокрылых и шерстокрылых. Группы протеутерий, давшие начало грызунам, повидимому стояли уже далеко от тех форм, которые были родоначальниками современных зайцеобразных. Одной из важнейших групп плацентарных, возникших от протеутерий, были древние копытные (отряд Condylarthra), сформировавшиеся в середине позднего мела. Освоив растительный корм и захватив уровень консументов I порядка, они смогли существенно повысить численность, широко распространиться по суше и дать обширную адаптивную радиацию. Из группы древних копытных развились такие ветви, как отряды трубкозубых, даманов, сирен, хоботных, парнокопытных, мозоленогих, непарнокопытных, китообразных и др. При этом даже в пределах отряда разные группы могли возникнуть от разных предковых форм (например, зубатые и усатые киты).
Становление крупнейшего отряда плацентарных — хищных (Carnivora) — шло от протеутерий или от очень ранних древних копытных при переходе последних к плотоядности. По общей организации определенную близость к своим палеогенным предкам имеют псовые и виверровые, тогда как кошачьи являются наиболее видоизмененными и специализированными, но сравнению с предковыми формами. Древние хищные, приспособившись к обитанию в воде, смогли дать начало группе ластоногих, среди которых ушастые тюлени, видимо, наименее изменились по сравнению с ранними формами ластоногих, живших на рубеже палеогена и неогена.
Таким образом, эволюция основных групп позвоночных во многих случаях шла независимыми, параллельными ветвями и не равномерно по времени и скорости. Такой «мозаичный» характер эволюции, без сомнения, отражается на строении организма животных, включая и их ЦНС, вследствие чего приходится безоговорочно отказаться от «линейных» схем эволюции как на уровне групп позвоночных животных разных рангов, так и на органном и тканевом уровнях организации животных.
- [1] Это явление в эволюции позвоночных было описано в трудах А. Н. Павлова (1901) как принцип филогенетического ускорения и предварения филогении онтогенией и в дальнейшем изучалось акад. Л. С. Бергом и его последователями, получив наименование «филогенетическоеопережение*. Принцип «филогенетического опережения* есть реальный результат неравномерности эволюционного процесса, характерной для всех уровней организации живого.
- [2] Необходимо подчеркнуть, что по современным палеонтологическим даннымкостные рыбы произошли независимо и одновременно с хрящевыми рыбами.
- [3] Это во многом определяет значительный интерес нейробиологов к изучению ЦНС черепах (как образно отмечено в одной из статей по морфологиимозга черепах — «ключа* к пониманию эволюции мозга млекопитающих).Это тем более интересно, поскольку существует точка зрения на происхождение черепах от основания эволюционной линии терапсид.
- [4] Отметим, что представителей отряда насекомоядных ни в коем случаенельзя рассматривать как предковые формы для других отрядов плацентарных (как это полагали ранее).