Строение митохондриального генома Saccharomycescerevisiae

Отдельные элементы регуляторной области генов, называемые энхансерами, могут располагаться перед структурной частью гена, позади нее или даже в ней самой. В структурной части большинства митохондриальныхгенов кодирующие последовательности нуклеотидов — экзоныперемежаются протяженными некодирующимипоследовательностями — интронами (рисунок 2). Суммарный размер интронов, как правило, многократно… Читать ещё >
Строение митохондриального генома Saccharomycescerevisiae (реферат, курсовая, диплом, контрольная)
Отдельные элементы регуляторной области генов, называемые энхансерами, могут располагаться перед структурной частью гена, позади нее или даже в ней самой. В структурной части большинства митохондриальныхгенов кодирующие последовательности нуклеотидов — экзоныперемежаются протяженными некодирующимипоследовательностями — интронами (рисунок 2). Суммарный размер интронов, как правило, многократно превышает общий размер экзонов гена. Исходя из этого факта, можно сделать вывод о том, что митохондриальный геном любого эукариотическогоорганизма содержит не только последовательность нуклеотидов с генетической информацией о белках и рибонуклеиновых кислотах (тРНК, рРНК и некоторых других), но и большоеколичество последовательностей нуклеотидов, не несущих информации.
Помимо интронов, в геноме Saccharomycescerevisiae имеется большое количество других некодирующих последовательностей нуклеотидов, главным образом различных повторяющихся последовательностей. Поэтому общая длина некодирующих последовательностей нуклеотидов в геноме Saccharomycescerevisiae в десятки раз превышает длину кодирующих последовательностей [2].

Рисунок 2. Гены цитохромаb и соответствующие мРНК у дрожжей и человека. Экзоны выделены цветом; интроны в дрожжевой ДНК выделены точками Митохондриальную ДНК S. Cerevisiae в редких случаях удавалось выделить в виде замкнутого кольца размером 25 мк. Как и в случае с мтДНК растений, обнаруживаемые линейные ДНК молекулы первоначально считали результатом разрывов ДНК invitro в процессе приготовления препаратов. Однако линейная структура молекул мтДНК обнаруживалась не только при электронномикроскопических исследованиях, но и при рестрикционном картировании геномов, пульс-электрофорезе. В настоящее время линейные молекулы мтДНК обнаружены почти у трети из 60 изученных видов дрожжей, а также у ряда других таксономических групп, включая водоросли, паразитических простейших, плесневые грибы, оомицеты. При изучении структуры линейныхмтДНК обнаружены гомогенные терминальные структуры, что позволило ввести термин «митохондриальныетеломеры».
Изученные к настоящему времени митохондриальные геномы дрожжей чрезвычайно изменчивы по размерам (от 17,3 т. п.н. у Schizosaccharomycespombe до 101 т. п.н. у Brettanomycescustersii) и порядку расположения генов, в то время как общее число генов изменяется незначительно. Основные причины столь существенных различий в размере геномов объясняются наличием или отсутствием интронов и изменчивостью межгенныхучастков-спейсеров [1, 10].
Общее количество ГЦ-пар в мтДНК дрожжей чрезвычайно мало-18%; молекула состоит из ГЦи АТ-богатых зон. Половину всей мтДНК составляют АТ-богатые последовательности, содержащие менее 5% ГЦ-пар — так называемые спейсеры. Два-три процента генома составляют короткие участки, богатые ГЦ (более 50%). Половина этих участков содержит сайт рестрикции Haelll и называется сайт-кластерами, остальные несут сайт Hpall и названы ГЦ-кластерами. Длина ГЦ-кластеров обычно 30−60 п.н., число их на митохондриальный геном может превышать 150, обнаруживаются они по всему геному, но преимущественно в межгенных участках. Наконец, в оставшейся части генома содержание ГЦ-пар около 26%-это участки, состоящие из собственно генов (Приложение А) представлена карта митохондриального генома Saccharomycescerevisiae, на которой показаны известные гены, точки начала репликации и их ориентация [4].
Размер митохондриального генома у разных видов рода Saccharomyces обычно колеблется от 85 т. п.н. (так называемые длинные геномы) до 78 т. п.н. (короткие геномы) и 74 т. п.н. (сверхкороткие геномы). У S. cerevisiae количество молекул мтДНК на гаплоидную клетку составляет в среднем 21−23. Этот вид имеет «длинный» геном — 85 т. п.н. [1,3].
Митохондриальный геном одного из примитивных видов дрожжей-S.uvarum оказался значительно меньшего размера-57 т. п.н. Соответственно у данного вида имеется меньше точек начала репликации (ori) — четыре вместо восьми; ГЦ-кластеров всего 50−60, в то время как у S. cerevisiae их 200. Учитывая, что все характерные для дрожжей митохондриальные гены присутствуют и у S. uvarum, сокращение размера молекулы мтДНК можно с уверенностью отнести за счёт некодирующих участков. Заметим, что у S. cerevisiae из восьми точек ori-активными являются только четыре (рисунок 3).
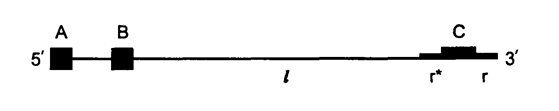
Рисунок 3. Схематичная структура ori последовательности. А, В, С — ГЦ-кластеры, r* и r — сайты, в которых репликация ДНК инициируется в обоих направлениях РНК-праймерами, l — АТ-богатый участок.
Ori-область митохондриальной ДНК дрожжей имеет обычно следующую структуру: три ГЦ-кластера, сайты инициации репликации и АТ-богатый участок длиной приблизительно 200 п.н. [4].