Адренорецепторы.
Строение и функции синапсов

Адренорецепторы являются гликопротеинами, ассоциированы с G-белками и имеют такое же строение, как и другие рецепторы этого типа. Их белковая цепь состоит из семи гидрофобных доменов в виде трансмембранной спирали, Домены соединены гидрофильными петлями, расположенными попеременно по обе стороны мембраны. вконец белковой молекулы адренорецептора расположен внеклеточно, С-конец — внутри клетки… Читать ещё >
Адренорецепторы. Строение и функции синапсов (реферат, курсовая, диплом, контрольная)
В 1948 г. английский фармаколог Рассел Алквист выдвинул гипотезу о двух типах адренорецепторов. аАдренорецепторы суживают сосуды, наиболее чувствительны к эпинефрину, меньше реагируют на норэпинефрин и очень слабо воспринимают действие изопреналина (изопропилнорадреналина). рАдренорецепторы расширяют сосуды, обладают максимальной чувствительностью к изопреналину, в 10−50 раз слабее реагируют на эпинефрин и норэпинефрин.
Адренорецепторы обнаружены на постсинаптической, пресинаптической мембранах и в клетках, не получающих адренергической иннервации. Постсинаптические адренорецепторы имеют индексы 1 или 2, пресинаптические и внесинаптические адренорецепторы обозначаются индексом 2. Внесинаптические адренорецепторы активируются циркулирующими в крови норадреналином и адреналином. рАдренорецепторы жировой ткани имеют индекс 3.
Все адренорецепторы характеризуются сходной последовательностью аминокислот (у а!- и а2- адренорецепторов идентичны 30% аминокислот, у рг и р2-адренорецепторов — 60%).
В сосудах и внутренних органах расположены аи р-адренорецепторы различных типов. Например, в сосудах легких обнаружено 30% ргадренорецепторов и 70% р,-адренорецепторов.
Адренорецепторы являются гликопротеинами, ассоциированы с G-белками и имеют такое же строение, как и другие рецепторы этого типа. Их белковая цепь состоит из семи гидрофобных доменов в виде трансмембранной спирали, Домены соединены гидрофильными петлями, расположенными попеременно по обе стороны мембраны. вконец белковой молекулы адренорецептора расположен внеклеточно, С-конец — внутри клетки. Активный центр адренорецептора представляет собой карман, образованный высококонсервативными аминокислотами, расположенными в средней и во внеклеточной третях гидрофобных трансмембранных спиралей. Аминогруппа катехоламинов соединяется ионной связью с карбоксилом аспарагиновой кислоты в третьем трансмембранном домене. Гидроксилы катехоламинов образуют водородную связь с остатками серина в пятом и седьмом доменах, что необходимо для активации адренорецепторов.
Сведения о механизмах функционирования, чувствительности к агонистам и антагонистам, физиологической роли адренорецепторов представлены в табл. 9−2-9−4.
Таблица 2. Адренорецепторы и их эффекторные системы.

а-Адренорецепторы
Постсинаптические а-|-адренорецепторы (типы А, В, D) активируют мембранные фосфолипазы и увеличивают проницаемость кальциевых каналов. В гладких мышцах ионы кальция активируют.
Примечания. Э — эпинефрин; НЭ — норэпинефрин; И — изопреналин.
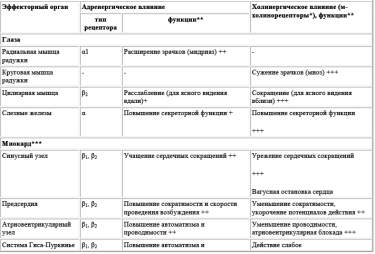


кальмодулинзависимую киназу легких цепей миозина, что необходимо для образования актомиозина и сокращения. Только в желудке и кишечнике а-|-адренорецепторы, открывая кальцийзависимые калиевые каналы, вызывают гиперполяризацию сарколеммы и расслабление гладких мышц. Эффекты активации аг адренорецепторов следующие:
- • сокращение радиальной мышцы радужки с расширением зрачков (мидриаз, от греч. ату^оэ — «темный, неясный»);
- • сужение сосудов кожи, слизистых оболочек, органов пищеварения, почек и головного мозга;
- • повышение АД;
- • сокращение капсулы селезенки с выбросом депонированной крови в циркуляторное русло;
- • сокращение сфинктеров пищеварительного тракта и мочевого пузыря;
- • подавление моторики и снижение тонуса желудка и кишечника. а2-Адренорецепторы (типы А, В, С) локализованы на постсинаптической, пресинаптической мембранах и вне синапсов.
Пресинаптические а2-адренорецепторы ингибируют аденилатциклазу и тормозят синтез цАМФ, увеличивают проницаемость мембран для К+ с развитием гиперполяризации, блокируют кальциевые каналы. По принципу отрицательной обратной связи они тормозят выделение норадреналина из адренергических окончаний при избыточной активации адренорецепторов.
Постсинаптические а2-адренорецепторы суживают сосуды кожи и слизистых оболочек, угнетают моторику желудка и кишечника, подавляют секрецию кишечного сока.
Внесинаптические а2-адренорецепторы суживают сосуды кожи и слизистых оболочек, угнетают моторику желудка и кишечника, секрецию инсулина, повышают агрегацию тромбоцитов.
в-Адренорецепторы
в-Адренорецепторы, активируя аденилатциклазу, вызывают превращение АТФ в цАМФ — активатор цАМФзависимых протеинкиназ. Протеинкиназа, А транспортируется в ядро клеток и фосфорилирует фактор транскрипции — ДНК-связывающий белок. Этот белок регулирует активность цАМФ-чувствительного элемента в промоторном участке гена, в результате активируется синтез в-адренорецепторов.
Для постсинаптических вгадренорецепторов характерны следующие эффекты:
- • стимуляция сердечной деятельности: тахикардия, ускорение проведения возбуждения по проводящей системе, усиление сокращений миокарда, увеличение потребности в кислороде (вгадренорецепторы при участии Эз-белков стимулируют фосфорилирование кальциевых каналов, что сопровождается их открытием, входом в саркоплазму Са2+ и мобилизацией этого иона из саркоплазматического ретикулума; также фосфорилируется белок фосфоламбан, способствующий депонированию Са2+ в саркоплазматическом ретикулуме);
- • стимуляция секреции ренина;
- • угнетение моторики кишечника.
Постсинаптические и внесинаптические в2-адренорецепторы расслабляют гладкие мышцы и вызывают гипергликемию. В гладких мышцах они при участии цАМФ снижают активность киназы легких цепей миозина, в печени и скелетных мышцах активируют фермент гликогенолиза фосфорилазу, ингибируют гликогенсинтазу. Кроме того, в гладких мышцах блокируются кальциевые каналы и активируются калиевые каналы. Типичные эффекты в2-адренорецепторов следующие:
- • расширение сосудов сердца, легких и скелетных мышц;
- • снижение АД;
- • расширение бронхов и угнетение секреторной функции бронхиальных желез;
- • угнетение моторики желудка и кишечника;
- • расслабление желчного пузыря, мочевого пузыря, матки;
- • усиление цАМФ-зависимого гликогенолиза и глюконеогенеза;
- • активация гликогенолиза в скелетных мышцах;
- • стимуляция секреции инсулина.
Пресинаптические р2-адренорецепторы осуществляют обратную связь, стимулируя выделение норадреналина при недостаточной активации адренорецепторов.
р3-Адренорецепторы усиливают цАМФ-зависимый липолиз в жировых депо с повышением в крови содержания свободных жирных кислот. Полиморфизм гена р3-адренорецептора создает предрасположенность к развитию ожирения и сахарного диабета 2-го типа.
р-Адренорецепторы подвергаются гомологической и гетерологической десенситизации. Гомологическая десенситизация развивается только к действию р-адреномиметиков. Под их влиянием медленно фосфорилируется специфическая цАМФ-независимая протеинкиназа — киназа р-адренорецепторов. Этот фермент фосфорилирует комплекс «р-адренорецепто-рЛ-адреномиметик». В дальнейшем присоединяется белок р-аррестин, нарушающий связь регуляторного домена р-адренорецепторов с G-белком. Инактивированный комплекс подвергается интернализации в цитоплазму и деградации в лизосомах.
При гетерологической десенситизации в ответ на действие р-адреномиметиков ослабляется реакция на многие вещества, повышающие синтез цАМФ. Она обусловлена быстрым (в течение миллисекунд) фосфорилированием р-адренорецептора протеинкиназой, А с последующей утратой способности передавать сигнал на эффекторные системы.