Структура и функции теломер клеток

Так как перемена условий обитания влечет за собой мобилизацию большинства функций в организме, то, несомненно, это приводит к стрессу и может влиять на процессы, связанные со старением. В этом отношении полезно вспомнить о теломерах, ведь эти концевые структуры хромосом могут служить удобной моделью отслеживания изменений, которые происходят в разные сезоны года, так как их основные свойства… Читать ещё >
Структура и функции теломер клеток (реферат, курсовая, диплом, контрольная)
Теломеры это концевые районы линейной хромосомной ДНК, состоящие из многократно повторяющихся коротких нуклеотидных последовательностей, сюда также входят многие белки, специфически связывающиеся с теломерными ДНК-повторами. Теломеры предотвращают изменение и слияние хромосом, поддерживая целостность генома клетки и ответственны за прикрепление хромосом к специальной внутриядерной структуре, и играют важную роль в создании специфической архитектуры и внутренней упорядоченности клеточного ядра. Сравнительно недавно молекулярными биологами были разработаны эффективные методы определения последовательности нуклеотидов в нуклеиновых кислотах. Первые исследования были проведены с одноклеточными простейшими из-за особенностей строения ядерного и хромосомного аппарата, содержащие несколько десятков тысяч мелких хромосом и, соответственно, множества теломер в одной клетке. Повторяющиеся много раз блоки в теломерной ДНК простейших состоят из шестивосьми нуклеотидных остатков, где одна цепь ДНК сильно обогащена остатками гуаниловой кислоты, а комплементарная ей цепь ДНК соответственно обогащена остатками цитидиловой кислоты.
У дрожжей повторяющиеся блоки намного длиннее, чем у простейших, но не столь регулярные. Учеными установлено, что теломерная ДНК человека, построенная из TTAGGG-блоков отличается от простейших лишь одной буквой в повторе. Более того, из TTAGGG-блоков построены теломерные ДНК всех млекопитающих, рептилий, амфибий, птиц и рыб, а в теломерной ДНК не закодировано никаких белков, а теломеры всех организмов выполняют универсальные функции. (Чернышев Н.В. 2010.).
К настоящему времени известно, что теломерные фрагменты у большинства живых существ очень схожи, они обогащены нуклеотидами G и С, которые располагаются в определенной, похожей у всех организмов последовательности. Несколько отличается структура ТR у некоторых видов грибов и особенно сильно — у дрозофилы. У Saccaromyces cerevisiae сравнительно короткие теломеры построены из нуклеотидов С и Т в последовательности, которая имеет непостоянную структуру Данные о строении теломер приведены в таблице 1.
Таблица 1 — Последовательность нуклеотидов теломер у разных видов живых организмов
Таксон. | Вид. | Последовательность нуклеотидов 5'>3'. | |
Простейшие. | Euplotes. | TTTTGGGG. | |
Слизневые грибы. | Phusarum. | TTTAGGG. | |
Жгутиковые. | Trypanosoma. | TTAGGG. | |
Споровики. | Plasmodium. | TT (T/C)AGGG. | |
Грибы. | Neurospora Candida maltosa. | TTAGGG ACGGATGGAGACTCGCTTGGTGT. | |
Нематоды. | Ascaris. | TTAGGC. | |
Насекомые. | Bombus mori. | TTAGG. | |
Водоросли. | Chlamidomonas. | TTTTAGGG. | |
Высшие растения. | Arabidopsis. | TTTAGGG. | |
Позвоночные животные. | Homo sapiens. | TTAGGG. | |
Кроме инфузории тетрахимены, теломерные повторы изучены у многих других представителей простейших, в том числе у стилонихии и окситрихи, у которых повторяющейся единицей является октамер 5'-ССССАААА-3'/ З'- GGGGТТТТ-З1. Этот октануклеотид расположен на концах хромосом особым образом, так что часть С-цепи ДНК остается одиночной из-за отсутствия второй цепи. Выступающие концы нитей ДНК обнаружены в теломерах всех организмов. Этот октануклеотид, выступающий за обычную двойную уотсон-криковскую спираль, называют одно цепочечным свободным G-концом. Длина простого повторенного дуплекса, расположенного проксимальнее, может варьировать от менее чем 50 пн (у Еир1оtеs) до более чем 100 тпн (у мыши). Интересно, что теломерные однонитевые свободные G-концы фактически одинаковы у многих представителей совершенно различных филогенетических таксонов, за исключением дрожжей. Анализ показал, что четыре молекулы гуанина на выступающем конце, не имеющие возможности контактировать с молекулами цитозина, как это бывает в обычной молекуле ДНК, все же спариваются. Они располагаются в одной плоскости и образуют водородные связи между собой.
Особенность формирования этой структуры заключается в том, что соединение нуклеотидов происходит не по принципу комплементарности оснований, а между молекулами одного и того же основания — гуанина.
Как уже отмечалось, рядом с теломерным повтором располагаются участки ДНК большой протяженности, называемые субтеломерными повторами — ТАS.
У дрожжей описано два субтеломерных повтора, X и Y'. X имеет размеры 0,3−3,75 тпн и менее консервативен, чем Y' (5,2−6,7 т п н). Они чередуются, или повтор может состоять только из Х-элементов. Последние имеют открытые рамки считывания. На основе слабой гомологии в первичных последовательностях с мобильными элементами предположили, что Y'- элементы возникли из одиночной инсерции в предковый геном дрожжей.(Канунго М. 1982.).
Субтеломерные повторы обнаружены у многих видов. Общая длина их может быть очень большой — у хирономуса до 300 тпн в каждой теломере.
По иронии судьбы термин «теломера» возник в результате исследований на дрозофиле. В то же время до сих пор нет никаких доказательств того, что в геноме дрозофилы есть короткие теломерные G-богатые последовательности, характерные для большинства других видов.
У дрозофилы на концах хромосом расположены теломеро-специфичные ретротранспозоны, НеТ-Аи ТАRТ. В общих чертах они имеют похожее строение. У обоих на 5'- конце присутствуют небольшие повторенные последовательности, а на З'-конце — более протяженный повтор, завершающийся длинным фрагментом, содержащим только нуклеотиды с аденином (А)n. Различаются они тем, что НеТ-Аимеет две частично наложенные одна на другую рамки считывания (ОRFs), кодирующие белки, в то время как ТАRТ ~ две, ОRF1 и ОRF2, расположенные тандемно. Оба рстротранспозона присутствуют во многих копиях, главным образом в прицентромерных районах. Они могут перемещаться по геному и в случае утраты теломерного района встраиваются на самый конец хромосомы, восстанавливая теломерную структуру (рис. 2).
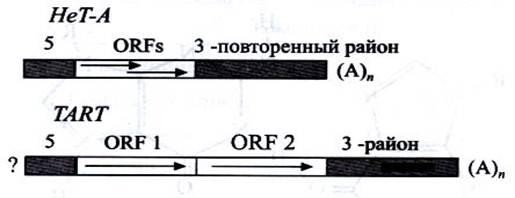
Рисунок 3. Строение теломер у дрозофилы.
Одной из важных характеристик теломерных ДНК является их длина. У человека она составляет от 2 до 20 тысяч пар оснований (т.п.о.), а у некоторых видов мышей может достигать сотен т.п.о.(Лэмб М.1980.).
Так как перемена условий обитания влечет за собой мобилизацию большинства функций в организме, то, несомненно, это приводит к стрессу и может влиять на процессы, связанные со старением. В этом отношении полезно вспомнить о теломерах [1], ведь эти концевые структуры хромосом могут служить удобной моделью отслеживания изменений, которые происходят в разные сезоны года, так как их основные свойства — динамичность и чувствительность к переменам во внешней и внутренней среде. Уже показано, что изменения в длине теломер связаны как со стрессом, так и со старением [2, 3, 4, 5]. И чем хуже внешние условия, тем нагляднее изменения в этих хромосомных структурах. В настоящее время существует достаточно работ, подтверждающих сезонную чувствительность теломер и их связь со стратегиями переживания или избегания неблагоприятных времен года.